Molecular Mechanisms of PCP Signaling
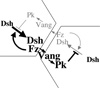
At the heart of PCP signaling is a mechanism for producing asymmetrically arrayed signaling components. The finding that distally localized Fz and Dsh designate the distal side of cells for prehair initiation raised the critical question of how these proteins achieve their asymmetric localization. We have proposed the following model to explain the Fz competition between adjacent surfaces of neighboring cells: Fz, Vang and Pk are initially uniformly distributed in the apical membrane, and Dsh is uniformly distributed in the cytoplasm. Fz recruits Dsh to form a DshFz complex, and, via Fmi homodimers spanning the intercellular junction, Fz also recruits Vang and Pk to the adjacent membrane of the neighboring cell. These five proteins therefore form a complex that spans neighboring cells, and these complexes are initially present in both orientations. However, both Pk and Vang cell-autonomously antagonize the DshFz interaction, thereby inhibiting formation of the oppositely oriented complex. The system is therefore inherently unstable in its starting state, and functions as a bistable switch, producing an all or none accumulation of complexes oriented in one direction. For several reasons, we propose that Pk and Vang antagonize Fz and Dsh localization by inducing endocytosis and removal of these components from a local region of the cell cortex. This feedback loop thus amplifies differences between Fz signaling levels on adjacent cell surfaces. The system relies on a globally distributed directional cue to push the switch in the correct direction.

Various data argue that the atypical cadherin Flamingo (Fmi) mediates communication between opposing membranes. We find that two functional forms of Fmi form asymmetric homodimers that signal instructively, producing a bidirectional and asymmetric signal that recruits Fz and Vang to opposite sides of the intercellular interface.
This model provides a satisfying explanation for many interesting phenomena in PCP signaling. However, a detailed molecular level understanding of this mechanism is lacking. We are examining several key questions, including how Pk and Vang mediate inhibition of Fz-Dsh association, how Fz and Vang communicate between opposing membranes, and how directional information feeds into this mechanism.

One source of global directional input is a module that serves to convert tissue level expression gradients to asymmetric subcellular Fat (Ft) - Dachsous (Ds) heterodimer localization, and provides directional information to the core module. It consists of the atypical cadherins Ft and Ds, that form heterodimers which may orient in either of two directions at cell-cell junctions, and the Golgi resident protein Four-jointed (Fj). Fj acts on both Ft and Ds as an ectokinase to make Ft a stronger ligand, and Ds a weaker ligand, for the other. As Fj and Ds are expressed in gradients across tissues, the result is the conversion of transcriptional gradients to subcellular gradients, producing a larger fraction of Ft-Ds heterodimers in one orientation relative to the other. Other less well defined sources of global directional information appear to act in partially overlapping, tissue dependent ways

A major question is to understand the relationship between the global directional cue and the core polarity amplificaiton and alignment mechanism. Our results suggest that the global system orients microtubules that in turn mediate directional trafficking of core PCP proteins to bias their localization. We are actively pursuing this hypothesis. Remarkably, the core PCP polarization mechanism differentially interprets the global signal, polarizing in opposite directions depending on the Prickle isoform present. We are studying how Prickle specifies this directional response.

We are using live imaging techniques to study the dynamics of PCP protein localization and further dissect the signaling pathway, and biochemical approaches to determine how PCP signaling proteins interact with the basic cell biological machinery.
An important goal in understanding development is to determine how distinct regulatory systems interact. In the Drosophila leg, mechanosensory bristles specify, via the EGF pathway, the formation of a cell known as a bract from the neighboring epidermal cells. In wildtype, the bract always forms proximal to the bristle, but in PCP mutants, this polarity is disrupted, as is the polarity of other structures within the epidermis. Therefore, the PCP system appears to hierarchically control polarization of a subsequent step in development. Other similar examples include the fly eye, in which PCP controls Notch signaling to determine R3/R4 photoreceptor cell fate, and fly leg joints, where PCP controls the polarity of joint orientation, probably also via Notch regulation. We are studying a variety of morphological processes in which PCP signaling specifies directionality.
People:
Maja Matis
Ying Peng
Gandhy Pierre-Louis
Jessica Oloffson
Katie Sharp
Alex Weiner
Bomsoo Cho